
Elu ja sümmeetria
Sümmeetria on üks looduse kõige nähtavamaid korraldusprintsiipe. Esmalt võib arvata, et loodus lausa armastab sümmeetriat. Bioloogias tähendab see kehaosade või elundite vastavust suuruse, kuju ja paiknemise poolest kujuteldavate sümmeetriatasandite või -telgede suhtes. Tegemist on lihtsa põhimõttega, mille taga peitub üllatavalt palju mitmekesisust.
Enamik hulkrakseid organisme on vähemal või rohkemal määral sümmeetrilised, kuid eluslooduses on sümmeetria alati ligikaudne. Näiteks taimelehed, mis on näiliselt sümmeetrilised, on harva keskelt kokkumurtuna täpselt ühesugused. Seega kasutab elu sümmeetriat ja asümmeetriat koos. Sümmeetria annab kehale korra, stabiilsuse ja äratuntava vormi ning asümmeetria teeb võimalikuks arengu, funktsioneerimise ja eristumise.
Sümmeetria kõige tuttavam näide on loomade bilateraalsümmeetria ehk kahekülgsus. Inimesel, linnul, kalal või putukal on eristunud ees- ja tagaots, kõhtmine ja selgmine pool ning vasak ja parem külg. Selline ehitus ei ole kaugeltki juhuslik. Bilateraalsus loob omamoodi ruumilise koordinaatsüsteemi, mille järgi arenevad organismi koed ja elundid. Evolutsiooniliselt oli bilateraalsuse teke suur samm, sest see võimaldas keerukamaid kehaplaane ja toetas suunatud liikumist.
Kõik elusolendid ei järgi siiski ühesugust mustrit. Paljud mereloomad, näiteks meduusid, merisiilikud ja meriroosid on radiaalse või radiaalsusele läheneva sümmeetriaga. Nende kehaosad paiknevad ümber keskse telje nii, et keha saab mitme tasandi järgi jagada sarnasteks või peaaegu sarnasteks osadeks. Selline ehitus on iseloomulik paiksetele või aeglaselt liikuvatele loomadele, kellele on kasulik tajuda ja mõjutada ümbritsevat keskkonda igast suunast enam-vähem võrdselt. Radiaalsümmeetria sobib hästi eluvormidele, kellel ei ole selgelt eristunud eesmist ja tagumist otsa, kuna nende eluviis eeldab keskpunktist lähtuvat organiseeritust. Loomulikult leidub ka organisme, kellel puudub igasugune sümmeetria. Sellisteks asümmeetrilisteks organismideks on näiteks amööbid ja käsnad.
Elule omaseks tunnuseks on veel ka see, et kuigi organismid võivad väljast paista sümmeetrilised, ei pruugi nad seda olla anatoomiliselt. Selgroogsed organismid, sealhulgas ka inimene, ilmestavad seda selgelt. Meie keha väliskuju on suuresti kahekülgne, kuid siseelundid ei paikne sugugi peegelpildis. Süda kaldub vasakule, maks on valdavalt paremal, soolestik ja teised elundid asetsevad aga hoopiski asümmeetriliselt. Selline vasak-parem asümmeetria ei ole kõrvalekalle normist, vaid normaalse arengu osa ning kujuneb embrüogeneesi käigus keerukate geneetiliste ja rakuliste protsesside tulemusena.1 Arengu käigus toimivad embrüos mehhanismid, mis määravad ära selle, kus on vasak ja kus parem pool. Pärast seda saab organism korraga säilitada teatud sümmeetrilised struktuurid, näiteks jäsemed või meeleelundite paarid ning samal ajal välja arendada vajaliku asümmeetria, mis puudutab peamiselt siseorganeid. Teisisõnu, normaalselt arenenud selgroogsel saab eristada nii sümmeetrilisi kui ka asümmeetrilisi osi.2
Hoopis veidralt tuleb eluslooduse sümmeetria esile molekulaarsel tasandil.
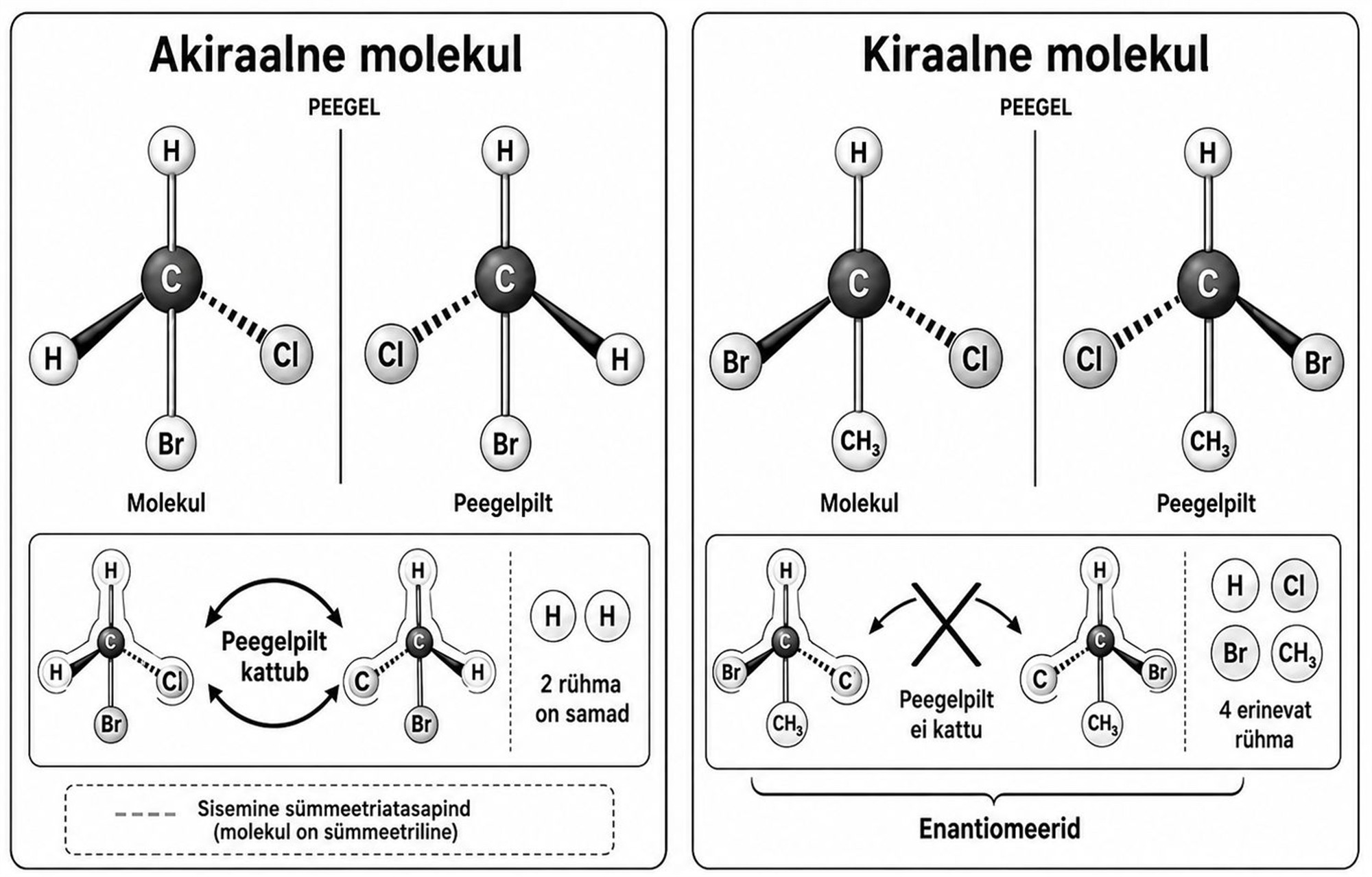
Peegelpildis molekulid
Molekulid ei erine üksteisest ainult selle poolest, millistest aatomitest nad koosnevad, vaid ka selle järgi, kuidas need aatomid on ruumis paigutunud. Mõnikord võib just see kolmemõõtmeline ehitus määrata, kuidas molekul käitub, milliste teiste molekulidega seondub ja millist rolli ta looduses mängib.
Molekulide ruumilise ehitusega on ka kiraalsus ehk käelisus. Kiraalsed molekulid on sellised, millel on kaks võimalikku vormi: need koosnevad küll samadest aatomitest, mis on seotud täpselt identsete keemiliste sidemetega, kuid nende ruumiline kuju on peegelpildis. Tavaliselt tuuakse siinkohal näiteks inimese vasak ja parem käsi või siis kindapaar, mida ei saa teineteise peale asetada nii, et kõik osad kattuksid. Selliseid molekulide peegelpildis vasteid nimetatakse enantiomeerideks.
Kiraalsus võib näida üksnes ebaolulise keemilise üksikasjana, kuid sellel on looduses määrav tähtsus – elusorganismide biokeemia on suures osas üles ehitatud just kiraalsetele molekulidele. Nii on käelised kõik aminohapped, millest koosnevad organismides leiduvad valgud. Aminohappel võib olla kaks peegelpildilist vormi: D (ld dexter, ’parem’) ja L (ld laevus, ’vasak’), mis viitab sellele, kuidas molekuli struktuur on seotud glütseraldehüüdi kahe peegelpildilise vormiga. Elusorganismides kasutatakse valkude ehitamiseks peaaegu alati L-aminohappeid. Erandiks on glütsiin, mis on liiga lihtsa ehitusega ega ole kiraalne. Samuti on kiraalsed DNA- ja RNA-molekulid tervikuna, kuid kiraalsed on ka nende olulised osad, eriti suhkrud riboos ja deoksüriboos.
Ka paljud teised organismides esinevad suhkrud, nt glükoos, on kiraalsed. Seejuures domineerivad eluslooduses enamasti D-suhkrud. Kõik rasvad ei ole kiraalsed, kuid paljud rakumembraani koostises olevad lipiidid jällegi on. Näiteks glütseroolil põhinevad fosfolipiidid võivad olla kiraalsed sõltuvalt sellest, millised rasvhapped on nendega seotud. Ka paljud väikesed metaboliidid ja signaalmolekulid, näiteks piimhape, aminohapete derivaadid, mitmed hormoonid, vitamiinid, terpeenid ja alkaloidid võivad olla kiraalsed.
Miks siis on kiraalsus oluline? Kõige üldisemalt võib öelda, et biomolekulide kuju määrab ära selle, kuidas nad üksteist ära tunnevad ja millistes reaktsioonides osalevad. Organismid ei kasuta biomolekulide kõiki võimalikke peegelpildis vorme võrdselt, vaid eelistavad väga kindlaid variante. See on üks elu kõige tähelepanuväärsemaid omadusi. Kõik tuntud eluvormid Maal kasutavad peaaegu eranditult vasakukäelisi aminohappeid ja paremakäelisi suhkruid (näiteks DNAs). Tegemist on omalaadse bioloogilise standardiga, mida nimetatakse homokiraalsuseks.
Homokiraalsuse päritolu
Kuidas homokiraalsus tekkis, on senini üks bioloogia ja astrofüüsika suurimaid mõistatusi. Teadlastel on selle selgitamiseks kolm peamist teooriat.
Tegemist võib olla lihtsalt juhusega, mis on evolutsiooni käigus kinnistunud. See tähendab, et elu kujunemisel tekkis esimene isepaljunev süsteem (näiteks RNA) puhtalt juhuse tahtel üht tüüpi kiraalsusega. Kuna organismis peavad kõik biokeemilised reaktsioonid omavahel ühilduma, hakkas see esimene edukas süsteem domineerima. Kui üht tüüpi kiraalsusega molekulid juba kasutuses olid, siis muutus hiljem nende vahetamine teist tüüpi molekulide vastu võimatuks, sest see oleks nõudnud kogu organismi üheaegset ümbertegemist. Tegemist on kõige lihtsama ja populaarsema teooriaga.
Teine teooria seostab homokiraalsuse tekke kosmiliste mõjudega. Üks põnevamaid leide seostub Murchisoni meteoriidiga, mis kukkus 1969. aastal Austraaliasse. Selles leidus rohkesti orgaanilisi ühendeid, sealhulgas aminohappeid. Need ei olnud pärit elusorganismidest, vaid olid tekkinud keemiliste protsesside tulemusena. Kõige üllatavam oli siiski see, et osal nendest aminohapetest ei esinenud L- ja D-vormid võrdses vahekorras. Tavaliselt tekib keemilistes reaktsioonides vasaku- ja paremakäelisi molekule täpselt ühesugune hulk. Meteoriidis oli aga vasakukäelistel aminohapetel kerge ülekaal. See vihjab millelegi väga olulisele, nimelt võis molekulaarne asümmeetria eksisteerida juba enne elu. Kuidas üldse saab elutu kosmos üht molekulaarset „kätt“ teisele eelistada? Üks võimalik vastus peitub valguses. Tähtede tekkepiirkondades, kus noorte hiidtähtede ümber keerlevad tolmu- ja gaasipilved, võib tekkida ringpolariseeritud valgus. Selline valgus võib peegelmolekulidega käituda veidi erinevalt. Üks molekuli vorm võib teatud tüüpi ringpolariseeritud valgust neelata pisut tugevamini kui teine. Kui valgus on piisavalt suure energiaga, võib see molekuli lõhkuda. Nii tekibki küll väike, kuid pika aja jooksul suuremaks saav erinevus L- ja D-molekulide vahekorras.
Kolmanda teooria järgi võib elu molekulaarne käelisus ulatuda lausa füüsika põhiseadustesse. Selle vaate pooldajate arvates on homokiraalsus seotud nõrga vastastikmõju ebasümmeetriaga. Nõrk vastastikmõju on üks neljast looduses esinevast põhijõust ja mängib näiteks tähtsat osa radioaktiivses lagunemises. Kui teised põhijõud on peegelsümmeetrilised, siis nõrk vastastikmõju on ainus fundamentaalne jõud, mis rikub paarsust. See tähendab, et osakeste nõrga interaktsiooni tulemused on erinevad, kui vaadeldakse protsessi peeglis. Seejuures eelistab nõrk jõud just vasakukäelist orientatsiooni. Teoreetiliselt on vasakukäelised aminohapped nõrga vastastikmõju tõttu imevähe stabiilsemad kui paremakäelised (vahe on umbes 10-14 eV). Kuigi see erinevus on üliväike, võis see ikkagi miljonite aastate jooksul toimides osutuda kaalukeeleks, mis suunas evolutsiooni eelistama vasakukäelisi aminohappeid.
See viib loomuliku küsimuseni, miks on elu kujunenud just ühe võimaliku lahenduse järgi, kuigi näib olemas olevat ka teine. Ja kas see teine variant on täielikult välistatud või lihtsalt seni teostumata jäänud võimalus?
Elu teisel pool peeglit
Peegelelu (ingl mirror life) oleks selline elu, mis põhineb paremakäelistel aminohapetel ja vasakukäelistel suhkrutel. Kui meie rakkude masinavärk koosneb üht tüüpi kiraalsetest molekulidest, siis peegelelu kasutaks nende peegelpildis vasteid. Sellistest peegelmolekulidest koosnevad peegelbakterid ei oleks veidi erinevad meile tuntud elust, vaid kuuluksid tervikuna hoopis teistsugusesse biokeemilisse maailma. Teadaolevalt looduslikku peegelelu olemas ei ole. Seega on tegemist hüpoteetilise võimalusega, mis on esile kerkinud koos sünteetilise bioloogia kiire arenguga. Inimesed suudavad sünteesida geene, genoome, valke ja mitmesuguseid rakulaadseid süsteeme, kuid täielikult autonoomset ja iseseisvalt paljunevat rakku ei ole seni suudetud veel luua. Kui peegelmolekulide loomine on laborites tavapärane, kuuluvad peegelorganismid esialgu veel ulmekirjandusse.
Bioloogiliste peegelmolekulide loomine on kiiresti arenev valdkond, mis pakub ka rakendusvõimalusi. Peegelpildilisi ehk D-aminohappeid saab tänapäeval valmistada mitmel viisil. Kõige lihtsamal juhul sünteesitakse korraga mõlemad vormid ja eraldatakse seejärel soovitud vorm. Täpsem ja moodsam meetod on juhtida reaktsiooni juba algusest peale nii, et peamiselt tekiks õige ruumilise kujuga ühend. Selleks kasutatakse spetsiaalseid katalüsaatoreid või ensüüme, mis „eelistavad“ üht peegelpildilist vormi teisele. Peegelpildilistest aminohapetest saab omakorda ehitada peegelvalke. Osa valke töötab organismides ensüümina, mis viivad läbi biokeemilisi reaktsioone. Tavapärased ensüümid ei suuda peegelmolekulidega reageerida, kuna need lihtsalt „ei mahu“ aktiivtsentrisse samamoodi nagu õige käelisusega ühendid. Selle takistuse ületamiseks on loodud peegelensüüme, mis reageerivad just nimelt peegelmolekulidega. Nii on saadud peegel-DNA polümeraase ja RNA polümeraase, mis suudavad töödelda peegel-DNAd ja peegel-RNAd. Eriti tähtis samm oli peegelpildilise T7 RNA-polümeraasi loomine, mis suudab valmistada täispikki peegelkujulisi ribosomaalse RNA molekule.3 Ribosoomid valmistavad aminohapetest valke. Kui õnnestuks valmistada täisfunktsionaalne peegelribosoom, peegelvalkude valmistamine lihtsustuks ja ühtlasi oleks astutud suur samm peegelelu loomise poole.
Kuna organismi tavalised ensüümid ei tunne neid hästi ära, võivad peegelmolekulid laguneda aeglasemalt, püsida kauem veres või kudedes ning kutsuda esile vähem soovimatuid immuunreaktsioone. Seega on neil perspektiivi ravimite ja diagnostiliste tööriistade arendamisel.
Miks peegelelu võib olla ohtlik?
Peamine mure seoses peegelorganismidega on seotud nende võimega hiilida mööda inimese ja teiste organismide immuunsüsteemist. Inimeste, loomade ja tõenäoliselt ka paljude teiste organismide kaitsemehhanismid tunnevad sissetungijad sageli ära just molekulide „käelisuse“ järgi. Kui bakter koosneks täielikult peegelpildilistest molekulidest, siis ei pruugi organismi kaitsesüsteemid seda ära tunda. Seega võivad sellised nakkused kujuneda väga raskesti kontrollitavaks ja isegi surmavaks.
Teine suur oht puudutab selliste peegelbakterite mõju olemasolevatele ökosüsteemidele. Bakterid on küll väikesed organismid, kuid neid esineb kõikjal suurel hulgal. Näiteks sisaldab 1 g mulda kuni 100 miljonit bakterit, aga kui me need üksteise taha rivistaksime, saaksime kõigest ühe sentimeetri pikkuse lõigu (bakterid on väga väikesed!). Bakteritel on siiski väga suur ökoloogiline tähtsus. Nende elutegevus mängib otsustavat osa näiteks nii süsiniku- kui ka lämmastikuringes. Kuna bakterid paljunevad pooldudes, võib nende hulk soodsates tingimustes kasvada hämmastava kiirusega. Tegemist on ju eksponentsiaalse kasvuga, mille käigus ühest bakterist saab kaks, seejärel kahest neli, neljast kaheksa jne. Kui bakter jaguneb iga poole tunni järel, võib ühestainsast rakust ööpäevaga saada üle 280 triljoni järglase. Kuigi üksik bakter on peaaegu kujuteldamatult väike, kaaluks selline hulk kokku juba umbes veerand kilogrammi. Looduses hoiavad baktereid vaos mitte ainult konkurendid, vaid ka bakteriofaagid, protistid, nematoodid ja teised „kiskjad“. Baktereid hävitavad organismid ei pruugi aga peegelbaktereid tuvastada ning need võivad keskkonnas laialt levida, häirida aineringeid ja ökosüsteeme.
Peegelbakteritega oleks ka väga raske võidelda, kuna nii nende tuvastamine kui ka hävitamine sõltuvad paljuski molekulide „käelisusest“. Mõned antibiootikumid võivad küll peegelbakteritele toimida, kuid isegi siis oleks nende globaalse leviku peatamine väga vaevaline. Nagu näha, on peegeleluga seotud riskitegurid: see on immuunsüsteemile raskemini äratuntav, olemasolevad looduslikud kontrollimehhanismid ei pruugi nendega toime tulla ja lisaks kõigele on tegemist isepaljuneva süsteemiga. Teadlaste hinnangul teebki just nende omaduste koosmõju peegelelust erakordselt tõsise bioloogilise ohuteguri.
Aga miks peaksime selle pärast üldse muretsema, kui peegelelu Maal ei leidu? Oht seisneb selles, et peegelelu võidakse luua laboris.

Kas peegelelu saab alguse laborist?
Algul näib see küsimus absurdne. Kuna peegeleluga seotud võimalikud ohud on küllatki hästi teada, siis miks peaks keegi proovima seda teha. Siinkohal tuleb mängu biotehnoloogia sisemine loogika. Kuna peegelmolekulidel on lai kasutusvaldkond, siis tegeletakse paljudes laborites nende valmistamise ja uurimisega. Selle tegevuse eesmärgiks ei olegi alternatiivse peegelelu loomine, vaid arendatakse püsivamaid ravimeid, täpsemaid diagnostilisi tööriistu ja vastupidavamaid bioloogilisi materjale. Iga selline ettevõtmine on küll igati põhjendatud ja kasulik, kuid koostoimes võivad need viia ohtliku tagajärjeni. Esialgu sünteesitakse üksikuid molekule, seejärel ühendatakse need keerukamateks struktuurideks ja seejärel hakatakse looma süsteeme, mis matkivad elu põhifunktsioone. Seetõttu ongi eksperdirühmad rõhutanud, et peegelmolekulide uurimine ja kasutamine on lubatav, kuid isepaljunevate peegelorganismide loomine peab olema täielikult keelatud.4
Arutelu peegelelu üle on mõneti erandlik. Tavaliselt hakatakse teaduslikke riske tõsiselt arutama alles siis, kui vastav tehnoloogia on juba käeulatuses või isegi kasutuses. Antud juhul püütakse aga peegelelu loomisega seonduvat reguleerida juba ennetavalt. 2024. aasta detsembris avaldas 38 teadlast maailma juhtivas loodusteaduste ajakirjas Science hoiatuse, et peegelbakterite loomine võib kaasa tuua ravimatuid nakkusi ja pöördumatut ökoloogilist kahju.5 2025. aastal toimus Pasteuri instituudis rahvusvaheline konverents, kus samuti jõuti järeldusele, et peegelorganismide loomisest tuleb hoiduda.6
Peegeleluga seotud arutluste taustal meenutatakse sageli nn Asilomari pretsedenti. 1970. aastate alguses, kui rekombinantse DNA tehnoloogia alles kujunes, tekkis teadlastel tõsine küsimus, kas erinevatest organismidest pärit DNA ühendamine võib põhjustada mingeid uusi ootamatuid probleeme. 1974. aastal kutsusid juhtivad molekulaarbioloogid üles kehtestama ajutine paus kõige riskantsematele sellelaadsetele katsetele.
Leiti, et vaja on hinnata selliste katsete võimalikke tagajärgi. Sellele järgnes 1975. aasta veebruaris Californias Asilomari konverentsikeskuses toimunud nõupidamine, kus püüti esimest korda süsteemi seada, millistel tingimustel tuleks uut tüüpi geneetilist uurimistööd jätkata. Konverentsil toodi välja keskse ideena, et võimalikke riske tuleb hinnata mitte tagantjärele, vaid juba eksperimentide kavandamise ajal. Need arutelud said otseseks eelkäijaks 1976. aasta Ameerika Ühendriikide riiklike terviseinstituutide (National Institutes of Health, NIH) juhistele, mis kujundasid rekombinantse DNA uurimise bio-ohutuse raamistiku aastakümneteks.7 Tehniliste ohutusreeglite kõrval osutus veelgi tähtsamaks see, et teadlaskond ise suutis kehtestada vastutustundliku eneseregulatsiooni. Just sellepärast meenutatakse Asilomari kohtumist tänapäeval ikka ja jälle, kui vaieldakse uute ja üha võimsamate tehnoloogiate ohutuse üle.8 Samal ajal tuleb tunnistada sedagi, et tollased arutelud keskendusid siiski peamiselt sellele, kuidas uut tehnoloogiat turvalisemalt kasutada ning küsimused, kelle huvides seda üldse arendatakse ja millised on tehnoloogia laiemad eetilised ja poliitilised tagajärjed, jäid tagaplaanile. Peegelelu korral on probleem märksa keerulisem, kuna küsimus ei ole ainult selles, kuidas tagada arendatava tehnoloogia turvalisus, vaid selles, kas mõni uurimissuund tulekski põhimõtteliselt keelata.
Peegelelu üle peetav debatt ei piirdu ainult selle, küll olulise, teemaga. Tegemist on probleemiga, mis üha enam kummitab tänapäeva teadust. Viimased sajandid on ühiskonnas kujundanud arusaama, et teadus teostab kõik varasemad unistused ja lahendab kõikvõimalikud inimkonna ette kerkivad probleemid. Tänapäeva teadus ei piirdu ainult teadmiste hankimisega, vaid ka nende üha kiirema ja ulatuslikuma tehnologiseerimisega. Eilne teooria on tänane laboriprotseduur ja juba homme rakendatav tehnoloogia.
Ühelt poolt nõuab ühiskond, et teadus annaks kiiresti kasutatavaid tulemusi, teiselt poolt tekitab see olukorra, kus tehniline võimalikkus hakkab iseennast õigustama. Kui midagi on võimalik teha, siis seda ka tehakse. Sünteetilise peegelelu loomise korral võib kaalul olla kogu senine eluslooduse korraldus ja tehtud vigu ei pruugi enam üldse olla võimalik tagantjärele heastada. Võib-olla on peegelelu kõige suurem õppetund just see, et mitte kõik võimalik ei pea saama tegelikuks. Ajaloos on piisavalt näiteid sellest, kuidas tehnilise võimekus jõudis inimesteni varem kui tarkus seda tehnoloogiat kasutada. Kas teaduse areng peab alati seisnema uute valdkondade hõivamises või võivad mõned piirkonnad jääda puutumatuks? Ennekõike peavad siinkohal peeglisse vaatama teadlased ise ja kaaluma võimalike tehnoloogiliste edusammude kõrval ka iseenda väärtusi, norme ja piire.
1 Ángel Raya, Juan Carlos Izpisúa Belmonte, Left–right asymmetry in the vertebrate embryo: from early information to higher-level integration. – Nature Reviews Genetics 2006, kd 7, lk 283–293.
2 Daniel T. Grimes, Making and breaking symmetry in development, growth and disease. – Development 2019, kd, 146, nr 16:dev170985.
3 Yuan Xu, Ting F. Zhu, Mirror-image T7 transcription of chirally inverted ribosomal and functional RNAs. – Science 2022, kd 378, nr 6618, lk 405-412.
4 Mirror life. Research and analysis. UK Government office of Science 16. VII 2025.
5 Katarzyna P. Adamala, Deepa Agashe, Yasmine Belkaid et al., Confronting risks of mirror life. – Science 2024, kd 386, nr 6728, lk 1351–1353.
6 Mirror life. Research and analysis. UK Government office of Science 16. VII 2025.
7 Donald S. Fredrickson, Asilomar and Recombinant DNA: The End of the Beginning. K.E. Hanna (toim), Biomedical Politics. National Academies Press, 1991, lk 258- 307.
8 Paul Berg, Asilomar, 1975: DNA modification secured. – Nature 2008, kd 455, lk 290–291.